|
|
Evolucion y mantenimiento del melanismo industrial en los Lepidoptera. Bol. SEA, 26, 1999 : 637-649 |
|
|
Evolución y mantenimiento del melanismo industrial en los Lepidoptera Michael E. N. Majerus
Introducción La evolución del melanismo industrial en los Lepidoptera, en general, y en la geómetra del abedul, Biston betularia, en particular, es el ejemplo más citado de evolución en acción. El reciente estudio de poblaciones naturales en Inglaterra y América ha descubierto que se han producido cambios evolutivos similares en diferentes subespecies de B. betularia (Fig. 1) en dos lugares independientes, lo cual ha proporcionando experimentos naturales replicados (Grant et al., 1996, 1998). En ambas poblaciones surgieron formas melánicas tras los cambios ambientales producidos por la contaminación industrial. La frecuencia de las formas melánicas alcanzó un máximo de un 90% alrededor de 1960 (Owen, 1961) y ha disminuido posteriormente a menos del 20%, tras la promulgación de la legislación contra la contaminación (Grant et al., 1996, 1998). La principal causa selectiva del incremento del melanismo en B. betularia, propuesta por primera vez por Tutt (1896) y confirmada por los experimentos clásicos de Kettlewell (1955a, 1956), fue la depredación diferencial por aves. En las regiones industriales, las formas melánicas se volvieron más crípticas que las no melánicas, en sus lugares de reposo sobre los árboles desprovistos de líquenes y tiznados de hollín, mientras que las formas no melánicas siguieron siendo más crípticas en las localidades rurales donde los árboles retenían una rica flora liquénica.
El caso de B. betularia ha sido tremendamente importante para la Ecología Evolutiva. No sólo proporciona uno de los mejores ejemplos observados de cambio evolutivo debido a selección natural, sino que ha demostrado que la selección natural puede ser una fuerza poderosa, ya que la forma melánica carbonaria debe tener 1,5 veces la eficacia biológica de la forma no melánica typica para dar cuenta de la rapidez de su auge inicial (Haldane, 1924). Además, ahora se ha demostrado que dicha evolución es reversible y replicable (Grant et al., 1996, 1998; Cook et al., en prensa). No obstante, incluso en B. betularia, el insecto con formas melánicas más estudiado, no se han desvelado por completo los detalles de los factores interactuantes que afectan a la evolución y el mantenimiento del polimorfismo melánico. Basándose en un modelo de eficacia biológica y migración (Mani, 1980), Many & Majerus (1993) han predicho que, en Gran Bretańa, carbonaria se extinguirá virtualmente en el ańo 2019, quedando reducida esta forma a la categoría de una mutación recurrente. Otros datos más recientes de Manchester, donde se registró por primera vez carbonaria en 1848 (Edleston, 1864), sugieren que esa predicción puede ser incluso demasiado optimista (Cook et al., en prensa). Parece, pues, oportuno revisar lo que se conoce y lo que se ignora sobre este caso, mientras existe aún alguna oportunidad de estudiar su declive. Biston betularia es sólo una de unas 200 especies británicas de Lepidoptera con formas melánicas, aunque sólo unas pocas muestran un melanismo de características similares (White, 1877; Kettlewell, 1973; Majerus, 1998). Aún así, el caso de B. betularia, en su formulación más simplista, se extrapola frecuentemente a otros Lepidoptera melánicos de Gran Bretańa y de otros lugares. Dicha extrapolación, basada en la depredación diferencial por aves en un ambiente cambiante, es inapropiada debido a que la evolución inicial del melanismo en muchos Lepidoptera no ha sido influida por la industrialización. En consecuencia, esta revisión comienza con una nueva clasificación del melanismo, adaptada de Kettlewell (1973). Después se considerará en detalle el caso de B. betularia, antes de discutir otros casos de melanismo industrial.
Una clasificación del melanismo en los Lepidoptera El melanismo en B. betularia es inusual porque constituye uno de los pocos ejemplos en los Lepidoptera en que las formas melánicas no estaban presentes en frecuencia apreciable antes de la revolución industrial. En la mayoría de las especies, las formas melánicas ya existían en algunas poblaciones antes de que las sustancias industriales contaminantes comenzasen a afectar de modo perceptible a los ambientes naturales. En algunas de estas especies, la frecuencia de las formas melánicas, y su oscuridad, han aumentado posteriormente en las regiones industriales. En otras, los melanismos han existido largo tiempo, y todavía existen en las poblaciones, independientemente del influjo de la industrialización. En consecuencia, pueden definirse tres grandes clases de melanismo en los Lepidoptera.
Melanismo industrial El melanismo industrial es el aumento proporcional de pigmentos melánicos en los miembros de una población, bien sea como resultado de un aumento en la frecuencia de distintas formas melánicas, bien como consecuencia de un oscurecimiento general de algunas o todas las formas dentro de una población, donde este aumento está asociado a los efectos de la contaminación industrial (Majerus, 1998). Se reconocen tres subclases de melanismo industrial: (1) polimorfismo melánico industrial completo, en el cual las formas melánicas han alcanzado frecuencias significativas sólo tras, y como consecuencia de, la industrialización, (2) polimorfismo melánico industrial parcial, en el que las formas melánicas presentes antes de la industrialización se han extendido geográficamente, o aumentado en frecuencia, desde, y como consecuencia de, la industrialización, y (3) melanismo industrial poligénico, en el cual el color medio de fondo de algunas poblaciones se ha vuelto gradualmente más oscuro como consecuencia de los efectos de la industrialización sobre el ambiente. En estas especies, se asume que el oscurecimiento del color de fondo es heredable y está controlado por cierto número de genes.
Melanismo no industrial en Lepidoptera crípticos Esta clase agrupa normalmente a Lepidoptera nocturnos que dependen de la cripsis para su defensa diurna, y en los cuales la existencia y frecuencia de formas melánicas es independiente de los efectos de la industrialización. Además, incluye el melanismo de algunos Lepidoptera diurnos. Si se hacen algunas modificaciones a la clasificación de Kettlewell (1973), pueden distinguirse ocho subclases, según las características geográficas de las poblaciones que muestran el melanismo, o los factores selectivos más probables para su explicación (Tabla 1). La comprensión de la evolución y la genética del melanismo no industrial puede resultar crucial para la investigación del melanismo industrial, ya que en muchos casos la fuente de formas melánicas industriales pueden haber sido melanismos que evolucionaron al margen de los efectos de la industrialización (Kettlewell, 1973; Majerus, 1998). A pesar de esto, se ignora el control genético del melanismo en muchas especies con melanismo no industrial. Además, sólo en un puńado de las numerosas especies implicadas se ha realizado trabajo experimental riguroso, en el campo y en el laboratorio, para descubrir o verificar las causas evolutivas del melanismo.
Melanismo en Lepidoptera con coloración de advertencia Kettlewell (1973) seńala que muchas especies diurnas de Lepidoptera que tienen coloración vistosa poseen formas melánicas. Se distinguen tres subclases: (1) melanismo recesivo, implica la existencia de formas melánicas en muchas especies aposemáticas, con un control de aquéllas por alelos recesivos raros, probablemente perjudiciales y mantenidos por mutación recurrente, (2) melanismo mimético, en el que especies comestibles poseen formas melánicas que son imitadoras batesianas de especies no comestibles, y (3) melanismo sexual, en el cual las formas melánicas han evolucionado como resultado de la selección sexual. Es improbable que exista una interacción evolutiva fuerte entre estas subclases de melanismo y el melanismo industrial en los Lepidoptera, aunque se conocen interacciones entre industrialización, melanismo térmico y otros factores selectivos en algunos Coleoptera aposemáticos, como Adalia bipunctata (Majerus, 1998). Para una discusión detallada del melanismo no industrial en los Lepidoptera crípticos y del melanismo en los Lepidoptera con coloración de advertencia, véase Kettlewell (1973) o Majerus (1998).
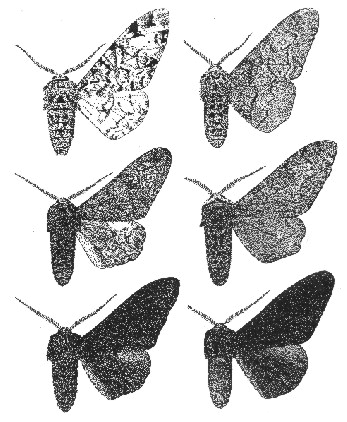
Polimorfismo melánico industrial completo El caso de B. betularia B. betularia es el ejemplo clásico de polimorfismo melánico industrial completo. Este caso se cita frecuentemente como el mejor ejemplo de 'evolución en acción'. Generalmente la historia que le cuenta, a menudo de modo muy breve, es que la forma negra de B. betularia surgió en el siglo XIX en partes industriales de Gran Bretańa, y aumentó su frecuencia debido a que estaba mejor camuflada en los árboles, desprovistos de líquenes y tiznados de hollín, que la forma no melánica. En consecuencia, era depredada con menor intensidad por las aves (Tutt, 1896), un punto de vista confirmado en los ańos 50 por el trabajo de Kettlewell en bosques de áreas contaminadas y no contaminadas (Kettlewell, 1955a, 1956). Las razones para la posición privilegiada de este ejemplo son fáciles de comprender. En primer lugar, los cambios en algunas poblaciones fueron rápidos y visualmente dramáticos. Una especie de polilla blanca y negra, relativamente invariable, se volvió totalmente negra en algunas poblaciones en un periodo de sólo 50 ańos. En segundo lugar, se observó que el cambio estaba asociado a un factor ambiental específico, el oscurecimiento de la corteza de los árboles debido a la contaminación atmosférica. En tercer lugar, un factor selectivo simple y fácil de entender, la depredación diferencial por aves, parecía suficiente para explicar el cambio evolutivo. Cuarto, la rapidez del aumento de la forma melánica indicaba diferencias selectivas de bastante mayor magnitud que las imaginadas por los primeros darvinistas. Por último, pudo presenciarse el cambio. Como escribió Sewall Wright (1978), el de B. betularia es: "el caso más claro en que se ha observado un proceso evolutivo conspicuo". En los 40 ańos transcurridos desde el trabajo pionero de Kettlewell, muchos biólogos evolutivos han estudiado el melanismo en B. betularia. Sus hallazgos muestran que el relato abreviado es inexacto o incompleto en lo que respecta a muchas de sus partes componentes. Cuando se consideran los detalles de la genética, el comportamiento y la ecología de la polilla, el panorama resultante es de mayor complejidad e interés que la simple historia que suele contarse. Los componentes de la historia normalmente mencionados figuran en la tabla 2, y se han considerado detalladamente en otros trabajos (Majerus, 1989). En este artículo, solamente se revisarán las variaciones importantes entre el relato abreviado y los hechos. Las formas de B. betularia y su herencia Biston betularia no tiene simplemente dos formas discretas. Existen formas intermedias entre typica y carbonaria, que comprenden un grupo de formas conocido colectivamente como insularia. El estudio acerca de las formas insularia ha sido comparativamente escaso, a pesar de que en muchas poblaciones la frecuencia de las formas insularia supera actualmente a la de carbonaria, y ha aumentado en algunos lugares a expensas de carbonaria tras la reducción de la contaminación (Brakefield, 1990; Majerus, 1998).
El complejo de formas insularia está controlado al menos por tres alelos adicionales del locus génico que controla carbonaria. Todos son más o menos recesivos frente a carbonaria y dominantes respecto a typica (Clarke & Sheppard, 1964; Less & Creed, 1977; Steward, 1977a), y su dominancia aumenta con su oscuridad (Fig. 2). Existe cierta evidencia de que en el siglo XIX la dominancia de carbonaria sobre typica era incompleta, y de que la selección natural actuó sobre modificadores que afectaban a la oscuridad de los heterocigotos en las fases iniciales del auge de carbonaria (Kettlewell, 1958a, 1973), de forma que la dominancia completa evolucionó del modo imaginado por Fisher (1930). Esta evidencia, que no carece de críticos (Bishop et al., 1978; Lees, 1981; Mikkola, 1984), se basa en observaciones de especímenes victorianos similares a los carbonaria modernos, pero con pequeńas manchas de escamas blancas en el centro de las alas. Dado que carbonaria está actualmente declinando en muchas poblaciones británicas, la selección natural puede estar actualmente reduciendo la dominancia genética de carbonaria sobre typica (Majerus, 1998). Sería de enorme valor el someter a análisis genético, mediante cruces controlados con especímenes typica tanto de origen industrial como no industrial, a individuos que se ajustasen a la descripción de los presuntos heterocigotos carbonaria victorianos de Kettlewell.
El comportamiento diurno de B. betularia Gran parte del trabajo experimental sobre la eficacia biológica relativa de las formas de B. betularia en hábitats naturales ha consistido en la colocación de mariposas vivas o muertas sobre troncos de árboles y en el registro de la proporción de cada forma que desaparecía por depredación de aves, bien observada o inferida (Kettlewell, 1955a, 1956; Bishop, 1972). Sin embargo, la exactitud cuantitativa de las estimas de eficacia biológica obtenidas de dicho modo es cuestionable debido a que la geómetra del abedul rara vez reposa durante el día en lugares expuestos de los troncos (Mikkola, 1979, 1984; Howlett & Majerus, 1987; Liebert & Brakefield, 1987). El mayor conjunto de datos conocido sobre los lugares de reposo de B. betularia silvestres encontradas en posiciones naturales es de sólo 54 polillas, recogido a lo largo de un periodo de 35 ańos (Tabla 3). Estos datos sugieren que la parte inferior de ramas y ramitas laterales en la copa son los sitios habituales de descanso, lo cual corrobora los resultados de los experimentos que han investigado las posiciones de reposo de machos (Mikkola, 1979, 1984) y hembras (Liebert & Brakefield, 1987) de B. betularia en cautividad, y confirma las propias observaciones de Kettlewell (1958b): "mientras realizaba sueltas a gran escala de ambas formas en la naturaleza, al alba, he podido observar en muchas ocasiones a esta especie en su posición de reposo habitual, que es la parte inferior de las ramas grandes de los árboles y, con menor frecuencia, sobre los troncos".
Si se asume que la eficacia biológica relativa de las formas de B. betularia depende de su cripsis, su lugar de reposo diurno es decisivo para la estimación de las diferencias de eficacia biológica entre formas. En regiones expuestas a escasa contaminación atmosférica, los líquenes foliosos aparecen principalmente sobre los lados menos sombreados y más húmedos de los troncos, superficies superiores de las ramas principales y, más generalmente, dentro de la copa (Liebert & Brakefield, 1987). También los líquenes crustáceos aparecen sobre dichas superficies, aunque dominan sobre los troncos y las superficies inferiores de las ramas principales. En áreas afectadas por altos niveles de contaminación casi no existen líquenes sobre los árboles y, normalmente, sólo aparece Lecanora conizaeoides, que también falta en los lugares más contaminados (Cook et al., 1990). Estos son los dos extremos. Los patrones de recolonización de los líquenes es pertinente en la actualidad, como resultado de la reducción en los niveles de contaminación. La recolonización inicial de los árboles, denudados por la contaminación pasada, se produce por el crecimiento de nuevas ramas en la copa, con baja contaminación residual y una intensidad luminosa comparativamente alta (Brakefield, 1987). Las últimas zonas recolonizadas de los árboles son los troncos, como resultado del lavado de las partículas contaminantes. En consecuencia, las estimaciones de eficacia biológica de cada forma pueden diferir notablemente dependiendo del lugar del árbol sobre el que se dispongan las polillas. En la mayor parte de los experimentos de depredación, las polillas se situaron sobre el tronco de un modo concreto, por lo común en la posición en que parecían más crípticas. Esta práctica puede criticarse por dos razones. En primer lugar, porque tales métodos hacen la asunción implícita de que las aves depredadoras ven lo mismo que los humanos y, en segundo lugar, porque asume que todas las polillas descansarán en lugares que maximizan su cripsis. La asunción de que las aves ven como los humanos es claramente falsa. Casi todas las aves tienen al menos cuatro tipos de conos visuales, uno de los cuales es sensible a la luz ultravioleta, a la que nosotros somos virtualmente ciegos (Chen & Goldsmith, 1986; Jacobs, 1992). En realidad, los análisis sugieren que las aves son más sensibles al ultravioleta que a la luz en la parte 'visible' del espectro electromagnético (400-700 nm), con una sensibilidad máxima alrededor de los 370 nm (Burkhardt & Maier, 1989). No sólo ven bien en el ultravioleta, sino que las aves pueden también discriminar diferentes longitudes de onda dentro de ese rango (Emmerton & Delius, 1980). Además, las aves tienen gotitas oleosas de tamańo variable frente a sus conos, que pueden actuar como filtros y les proporcionan mayor capacidad de discriminación de los colores que la que nosotros tenemos (Partridge, 1989). Aunque todavía se ignoran muchos de los detalles de la visión de las aves, resulta claro que tienen una mayor agudeza visual que los humanos. Esto significa que deben reevaluarse los estudios de depredación de B. betularia por aves, diseńados por humanos con la asunción de que las aves ven como nosotros. Parte del razonamiento subyacente a la práctica de situar polillas en posiciones que maximizan su cripsis es la idea de que las formas de B. betularia se comportan de modo diferente, y que cada forma adopta la posición que maximiza su propio camuflaje. Así, carbonaria debería reposar preferentemente sobre fondos oscuros homogéneos, mientras que typica debería haber retenido un comportamiento ancestral de reposo sobre fondos más claros y heterogéneos. Utilizando polillas de origen desconocido y barriles forrados con superficies iguales de cartón blanco y negro, Kettlewell (1955b) demostró que typica reposaba preferentemente sobre las superficies blancas, y que carbonaria prefería las negras. Algunos estudios han obtenido resultados similares (Boardman et al., 1974; Kettlewell & Conn, 1977), pero no otros (Mikkola, 1984; Howlett & Majerus, 1987; Jones, 1993). Majerus (1989) ha revisado estos resultados contradictorios. El mecanismo de conflicto-contraste de Kettlewell (1955b) para la selección de sustrato debe ser rechazado ante la demostración, por Grant & Howlett (1988), de variación interfamiliar en dichas preferencias. También resulta improbable una explicación genética de la selección de sustrato en B. betularia específica de cada forma, basada en el ligamiento entre los alelos morfológicos y etológicos apropiados, debido al poco tiempo disponible desde la aparición de carbonaria para que la selección natural haya promovido dicho ligamiento. En su lugar, Majerus (1989) propuso que, en aquellos sitios donde carbonaria ha conseguido mayor frecuencia, se ha extendido una mutación que indujo a las polillas a reposar sobre fondos oscuros, de la cual se beneficiaría la mayor parte de la población. A la inversa, en las poblaciones en las que carbonaria permaneció escasa, las polillas han retenido el comportamiento ancestral de reposo sobre sustratos claros. Así, se pueden imaginar tres tipos de poblaciones: (1) tipo a, en las cuales nunca se ha extendido un alelo de preferencia por sustratos oscuros, (2) tipo b, en las cuales dicho alelo ha surgido y constituye un polimorfismo transitorio a medida que se extiende hacia su fijación o eliminación, según el aumento o descenso de la frecuencia de carbonaria; en estas poblaciones algunas polillas preferirán sustratos claros y otras oscuros, pero las preferencias no estarán demasiado ligadas a ninguna forma, y (3) tipo c, en las cuales el alelo para la preferencia de sustratos oscuros se ha fijado y todas las polillas prefieren superficies oscuras. La evidencia en apoyo de esta hipótesis procede de pruebas que muestran diferencias entre poblaciones en el grado de preferencia por fondos claros u oscuros, y en las que las poblaciones de regiones industriales generalmente muestran mayores preferencias por sustratos oscuros, al tiempo que no muestran ninguna evidencia de preferencias de sustrato típicas de cada forma (Jones, 1993). Majerus (1989) argumenta que la preferencia observada de typica por lugares de descanso blancos y de carbonaria por lugares negros puede explicarse si Kettlewell (1955b) hubiese utilizado polillas de diferentes poblaciones, por ejemplo typica de poblaciones rurales (tipo a) y carbonaria de poblaciones industriales (tipo c). La misma teoría pudiera explicar otros resultados aparentemente contradictorios (Majerus, 1998). Las preferencias de sustrato son cruciales para interpretar otros resultados experimentales en B. betularia. Por ejemplo, los niveles de eliminación selectiva de typica en Birmingham y de carbonaria en Dorset en los experimentos de marcaje y recaptura de Kettlewell (1956) serían artificiales si las polillas liberadas procediesen de poblaciones con diferente preferencia en el lugar de reposo. Dado que Kettlewell observó que carbonaria estaba ausente de su lugar experimental en Dorset, las carbonaria que él utilizó allí deben haber procedido de otro lugar. Una tercera crítica va dirigida a los investigadores que, para sus experimentos de depredación, colocan polillas muertas en posiciones 'naturales' en lugar de utilizar polillas capaces de seleccionar su propia ubicación, como hizo Kettlewell (1955a, 1956). Estos investigadores asumen que su juicio es tan bueno como el de una polilla cuyo comportamiento ha sido moldeado por selección natural durante milenios, y para la cual una decisión errónea puede resultar letal. Sería muy conveniente la realización de experimentos para evaluar el grado relativo de cripsis de las polillas muertas situadas sobre troncos en posiciones 'naturales' y el de polillas vivas con posibilidad de elegir su lugar de reposo.
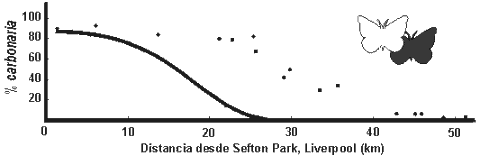
La dinámica geográfica de la frecuencia de melanismos La reciprocidad de los resultados de Kettlewell (1955a, 1956) en Birmingham y en Dorset proporciona evidencias sólidas de que la eficacia biológica de las formas de B. betularia difiere en ambientes contaminados y no contaminados. Otros estudios han demostrado diferencias de eficacia biológica similares (para una revisión véase Kettlewell, 1973; Lees, 1981; Majerus, 1998). En uno de los estudios más amplios, Bishop (1972) estudió un transecto desde el Sudoeste de Merseyside al Norte de Gales. Cuantificó la eficacia biológica relativa de las formas a lo largo de esa clina, sobre la base de sus conocimientos de la ecología y la genética, y utilizó un modelo de ordenador que incorporaba toda la información obtenida para simular el auge y la dispersión de carbonaria a lo largo del transecto desde su aparición al comienzo del siglo XIX (Fig. 3). Después comparó la posición predicha por el modelo para la clina en 1970 con la frecuencia observada de las formas a lo largo del transecto. La correspondencia entre la frecuencia observada y simulada no fue buena (Fig. 4), ya que aunque el modelo predijo que la frecuencia de carbonaria disminuiría hacia el Sudoeste de Liverpool, la posición predicha de dicho descenso estaba más próxima a Liverpool de lo observado. Además, la tasa de declive en los datos observados fue más rápida de lo predicho.
El ajuste entre las frecuencias simuladas y observadas para este transecto puede mejorarse significativamente si se introducen grandes ventajas no visuales para carbonaria (Mani, 1980, 1982). Sin embargo, la pobreza del ajuste de la simulación de Bishop a las frecuencias observadas también puede deberse a errores cuantitativos en la eficacia biológica relativa obtenida a partir de experimentos de depredación, debido a que las polillas fueron situadas sobre los troncos, no bajo las ramas laterales o ramillas, donde es probable que, en la sombra intensa, carbonaria sea más críptica y goce de mayor eficacia biológica (Howlett & Majerus, 1987). Los resultados de los experimentos de depredación en bosques de áreas contaminadas y no contaminadas confirman esta opinión, aunque el conjunto de datos es pequeńo (Howlett & Majerus, 1987). Si esto se confirma, puede explicar parte de la falta de ajuste entre la frecuencia simulada por Bishop y la observada, ya que el efecto será un aumento de la eficacia biológica relativa de carbonaria a lo largo de la clina, lo cual empujará apreciablemente hacia el Este la posición predicha del descenso en la frecuencia de carbonaria. Majerus (1998) seńala además que la evolución de una preferencia por los sustratos oscuros en poblaciones industriales, frente a las rurales, puede explicar la otra discordancia entre la simulación de Bishop (1972) y la frecuencia observada. Si han evolucionado preferencias específicas de cada población, las formas más raras en cada extremo de la clina podrían tener preferencias de sustrato inapropiadas para su color y sufrirían mayor eliminación selectiva, lo cual agudizaría la clina.
La frecuencia de carbonaria nunca ha alcanzado el 100% en ninguna población estudiada. Esto se atribuye a dos factores. En primer lugar, dado que typica es recesiva frente a carbonaria, la erradicación final del alelo para typica será lenta incluso cuando la selección natural en su contra sea alta, puesto que los alelos typica quedarán ocultos a los efectos de la selección dentro de los heterocigotos. En segundo lugar, los individuos migradores que portan el alelo typica invadirán las regiones industriales desde otras menos contaminadas. En apoyo de la importancia de la migración en B. betularia, existe evidencia circunstancial procedente de comparaciones entre las distribuciones de frecuencia de formas melánicas de ésta y otras dos especies, Gonodontis bidentata y Phigalia pilosaria. De estas especies, B. betularia se considera la más dispersiva y G. bidentata la más sedentaria. Los cambios en la frecuencia melánica a escala geográfica son más rápidos en G. bidentata, intermedios en P. pilosaria y más graduales en B. betularia, como se esperaría de acuerdo con modelos de equilibrio selección-migración (Bishop et al., 1978; Conroy & Bishop, 1980; Lees, 1981). Aunque circunstancial, esta evidencia indica que la migración ejerce una influencia importante sobre la frecuencia de las formas melánicas en diferentes regiones. No obstante, debe seńalarse que el conocimiento de las tasas de migración en todas estas especies se basa en datos incompletos, y casi exclusivamente en experimentos de marcaje y recaptura con machos adultos. Como afirma Brakefield (1987), basar las tasas de migración simplemente en el vuelo de los adultos puede haber conducido a serias subestimaciones ya que las larvas de primer instar, suspendidas en hilos de seda, pueden dispersarse a largas distancias (Kettlewell, 1973). En realidad, la velocidad de movimiento de carbonaria en su radiación desde Manchester tras 1848 sugiere tasas de migración ocasionalmente superiores a la de 2 km por generación, normalmente citada (véase, no obstante, Brakefield & Liebert, 1990). Por ejemplo, en sólo 49 ańos, en el siglo XIX, carbonaria se extendió unos 300 km desde Manchester al New Forest (Majerus, 1998).
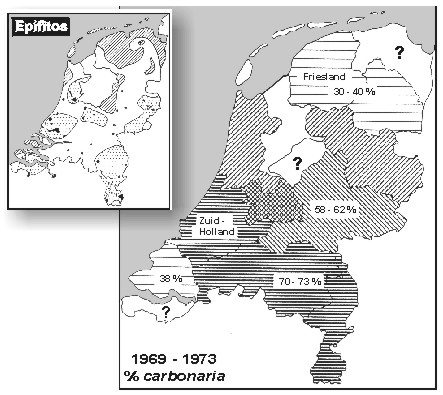
Melanismo en B. betularia fuera de Gran Bretańa El polimorfismo melánico en B. betularia no está limitado a Gran Bretańa. En Holanda, carbonaria fue registrada por primera vez en 1867 (Heylaerts, 1870) y estaba extendida en 1900, al igual que todo el conjunto de formas insularia (Brakefield, 1990). La comparación de la frecuencia de carbonaria con la industrialización y la flora epifita muestra relaciones similares a las de Gran Bretańa (Brakefield, 1990; Fig. 5). Sin embargo, la frecuencia de carbonaria en Holanda no ha alcanzado los máximos niveles registrados en Gran Bretańa, pues nunca ha superado el 75%, e insularia es generalmente más común en Holanda que en Gran Bretańa. En Luxemburgo, durante los ańos 70, carbonaria suponía un 70% de las poblaciones en los distritos industriales del Norte, pero sólo el 30% en Las Ardenas. En 1989-1993, la frecuencia de carbonaria ha disminuido significativamente en ambas regiones (Majerus, 1998). Tanto carbonaria como insularia son comunes en regiones industriales de Bélgica y del Norte de Francia, y en Alemania carbonaria ya era común en las regiones industriales en 1900 (Kettlewell, 1973). También se han documentado altas frecuencias de formas melánicas en regiones industriales de Dinamarca (Hoffmeyer, 1949), Suecia, Finlandia (Kettlewell, 1973) y Checoslovaquia (Novak & Spitzer, 1986). En Checoslovaquia, Novak & Spitzer (1986) demostraron una relación positiva significativa entre la frecuencia de carbonaria y el grado de contaminación atmosférica.
El melanismo industrial también se ha registrado en la subespecie americana, B. betularia cognataria. La forma melánica completa, swettaria, es indistinguible de carbonaria y también se hereda como un dominante unifactorial (West, 1977). La forma swettaria se encontró por primera vez en 1906 cerca de Filadelfia (Owen, 1961). En algunas partes de Michigan, la frecuencia de swettaria aumentó hasta un 90% en 1960. La forma swettaria ha sido hallada en otros puntos de Norteamérica, tales como Illinois, Massachusetts, Delaware, Montreal, Toronto y el Sur de Ontario (Owen, 1961; Kettlewell, 1973; Sargent, 1974; West, 1977). Existe una tercera subespecie, B. betularia parva, en Japón. El melanismo no ha evolucionado en dicha subespecie porque no aparece en regiones afectadas de modo importante por la contaminación industrial (Asami & Grant, 1995).
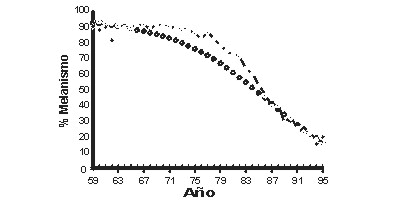
El declive del melanismo en la geómetra del abedul El declive actual de carbonaria en Gran Bretańa ha seguido al Acta del Aire Limpio de los 50, y a las leyes anticontaminación posteriores. A diferencia del auge de carbonaria, este descenso ha sido bien documentado (Cook et al., 1970, en prensa; Whittle et al., 1976; Howlett & Majerus, 1987; Mani & Majerus, 1993; Grant et al., 1995, 1996, 1998). La frecuencia anual de carbonaria entre 1959 y 1995, para una localidad en The Wirral, Merseyside, se muestra en la figura 6. El patrón de disminución coincide con lo esperado, ya que existe un retraso en el comienzo del declive de carbonaria tras la reducción en los niveles de contaminación. Se interpreta que dicho retraso representa el periodo necesario para el aclarado de las superficies de los árboles, por crecimiento de nuevas ramas no tiznadas, por lavado del hollín y por recolonización de los epifitos.
En Holanda y en los Estados Unidos se han registrado disminuciones similares. En Holanda, carbonaria está siendo reemplazada tanto por typica como por la más oscura de las formas insularia (Brakefield, 1990). En Michigan, el descenso en la frecuencia de swettaria refleja fielmente el descenso de The Wirral. En ambos casos, dicha disminución ha ido desde más del 90% a menos del 20% (Grant et al., 1996; Fig. 6). Aunque se carece de datos globales, se conocen declives similares en la frecuencia de formas melánicas en otras partes de América (Grant et al., 1998). Grant y sus colaboradores informan de que, en ambas regiones, la caída en la frecuencia de formas melánicas se relaciona bien con el descenso en SO2 y partículas en suspensión. Comentan que en ninguna localidad parece haber habido un aumento apreciable de la flora liquénica en ese periodo. Sin embargo, dado que las evaluaciones de los líquenes fueron anecdóticas, más que sistemáticas, y que pueden no haberse examinado las copas, este comentario debería considerarse provisional. Más aún, cuando otros autores han registrado un aumento de los líquenes tras la legislación contra la contaminación (Hawksworth & McManus, 1989; Bates et al., 1990; Cook et al., 1990). Las observaciones y experimentos de Kettlewell sobre B. betularia tuvieron una influencia considerable en la Biología Evolutiva, ya que movieron el principal mecanismo de evolución biológica desde la teoría al reino de los hechos. La reciente inversión en la fortuna de carbonaria ha brindado la oportunidad de cubrir un hueco importante en el conocimiento científico. Aunque se dice a menudo de B. betularia que hemos sido testigos de la evolución del melanismo, de hecho existen muy pocos datos precisos del aumento en frecuencia de carbonaria. En el momento en que los científicos comenzaron a examinar el caso en detalle, ya había ocurrido la mayor parte del incremento. El declive de carbonaria tras las leyes de aire limpio ha planteado la oportunidad de seguir de cerca el cambio de frecuencia y evaluar si los cambios observados pueden explicarse plenamente sobre la base de la comprensión actual del comportamiento y ecología de esta polilla.
Otros casos de polimorfismo melánico industrial completo Aparte de B. betularia, la cantidad de ejemplos de polimorfismo melánico industrial completo es escasa. Entre ellos se incluyen especies en las cuales se han descrito formas melánicas a partir de individuos aislados recogidos en regiones no industriales antes del establecimiento de auténticos polimorfismos genéticos (sensu Ford, 1940) (Majerus, 1998). En unas pocas especies el melanismo industrial se ha desarrollado muy recientemente, en los últimos 40 ańos, como en los casos de Brachionycha sphinx y Xylocampa areola. Las razones por las que el melanismo industrial no se desarrolló antes en dichas especies es, probablemente, azaroso: la mutación adecuada no surgió previamente en el lugar y momento correctos. Estos casos recientes de melanismo industrial subrayan que la selección natural no puede causar cambios excepto que disponga de variación fenotípica sobre la cual actuar. El ejemplo del pariente británico más próximo de B. betularia, Biston strataria, es muy instructivo en este punto. Esta especie tiene una forma melánica, melanaria, que es un melanismo industrial común en Holanda, pero nunca ha aparecido en frecuencia apreciable en Gran Bretańa. La ecología y el comportamiento de B. strataria son similares a los de la geómetra del abedul. Sin embargo, melanaria parece no haber surgido nunca en Gran Bretańa en circunstancias favorables y, por tanto, nunca se ha establecido allí. Al menos en una especie, el melanismo industrial británico es producto de la migración desde la Europa continental. La forma melánica, fusca, de Tethea ocularis apareció en el Sur de Inglaterra a mediados de los 40 y ha prosperado rápidamente desde entonces. El grupo de melanismos industriales completos también incluye formas fuertemente melánicas de algunas especies que previamente tenían formas melánicas menos extremas, de origen no industrial. Por ejemplo, Peribatodes rhomboidaria tenía una forma melánica no industrial, perfumaria, que aumentó su frecuencia alrededor de las conurbaciones. Sin embargo, en algunas localidades esta forma ha sido parcial o totalmente reemplazada por una nueva forma más extrema, rebeli. El declive en la frecuencia de formas melánicas en Gran Bretańa a lo largo de los últimos 30 ańos no se ha limitado a B. betularia. Aunque los datos publicados son escasos, la frecuencia de melanismos en diversas especies va actualmente en descenso (por ejemplo, West, 1992). La actual disminución de melanismos industriales completos en Gran Bretańa, Holanda, EE.UU. y, sin duda, en otras partes del mundo desarrollado no es general. Por ejemplo, en Finlandia Mikkola (1989) informó sobre una nueva forma melánica, inferna, de Xestia gelida, que él interpreta como el primer caso de un melanismo industrial en una zona subártica. Su aparición está asociada a la intensa contaminación procedente de la Península de Kola. Es probable que se descubran nuevos casos en países que sólo ahora están desarrollando una industria pesada y, con el declive de los melanismos industriales en Gran Bretańa, el estudio del melanismo industrial tendrá forzosamente que moverse a otra parte.
Polimorfismo melánico industrial parcial Esta categoría incluye a la mayoría de las especies que tenían formas melánicas antes de la revolución industrial, si su distribución incluye grandes regiones industriales. El caso de P. rhomboidaria, mencionado anteriormente, ilustra este punto. Originalmente un melanismo no industrial, perfumaria aumentó rápidamente su frecuencia en las regiones industriales a finales del siglo XIX (Jones, 1993). Casos similares incluyen a P. pilosaria, Allophyes oxyacanthae, Apanea crenata y Deileptenia ribeata, así como a ciertas especies que muestran preferencias de hábitat características de cada forma. Este último grupo de especies es de particular interés. La existencia de preferencias de hábitat típicas de cada forma en los Lepidoptera con polimorfismo melánico fue sugerida por vez primera por Kearns & Majerus (1987) para explicar las diferencias significativas en la frecuencia de Alcis repandata y Semiothisa liturata melánicos en capturas mediante trampas para polillas a lo largo de una distancia de menos de 20 m a cada lado de un límite brusco entre hábitats. Se han registrado posteriormente diferencias similares en otras 12 especies, con mayor frecuencia, en todos los casos, de formas melánicas en bosques con doseles densos respecto a hábitats adyacentes menos densos (Aldridge et al., 1993; Jones et al., 1993; Fraiers et al., 1994; Majerus et al., 1994; Majerus 1998). Kettlewell (1973) argumenta que muchas especies que actualmente exhiben melanismo industrial evolucionaron dicho melanismo antes de la industrialización. Majerus (1998), en concordancia con dicha opinión, sostiene que en el pasado distante, las formas melánicas evolucionaron en circunstancias ecológicas específicas, como en bosques de coníferas. Estas formas melánicas estaban en desventaja selectiva si abandonaban esas áreas con las características ecológicas especiales a las que estaban adaptadas. En consecuencia, los melanismos evolucionaron comportamientos que les restringían a dichos hábitats. Los cambios recientes en el manejo forestal, el uso de la tierra y el aumento de la contaminación han proporcionado nuevos hábitats (por ejemplo, plantaciones de coníferas, bosques contaminados) en los que las condiciones ecológicas favorecen a los melanismos y, en consecuencia, se han extendido y aumentado en frecuencia, al tiempo que retienen las características de hábitat específicas de su forma. Muchas especies que muestran melanismos para evitar las imágenes de búsqueda (Tabla 1) también tienen mayor frecuencia de formas melánicas en regiones industriales. Esto concuerda con lo esperado debido a que la probabilidad de que un depredador forme una imagen de búsqueda para un tipo particular de presa depende de la abundancia aparente de la presa para el ave que, a su vez, es una función de la abundancia y la cripsis. Muy pocas de las especies que caen dentro de esta clase de melanismo han sido estudiadas en profundidad. Las excepciones son P. pilosaria (Lees, 1981), A. repandata (Kettlewell, 1973; Jones et al., 1993) y A. oxyacanthae (Steward, 1977b). Todas ellas reposan sobre los troncos y el aumento de las formas melánicas en las regiones industriales se atribuye a su mayor cripsis.
Melanismo industrial poligénico De todas las clases de melanismo, el melanismo industrial poligénico es el menos estudiado y , quizá, el más difícil de estudiar. El estudio de material recogido a lo largo de los últimos 150 ańos revela un aparente oscurecimiento del color y la pérdida del diseńo en muchas especies de regiones industriales, independientemente de la forma a la que pertenezcan. Parte se debe a la decoloración de los ejemplares de museo, aunque es difícil achacar todas las diferencias a este único factor. La comparación de series de especímenes de seis especies, de regiones rurales e industriales, recogidos hace unos 80 ańos con los recogidos en los últimos cuatro ańos mostró que el color de fondo se había oscurecido más en las regiones industriales que en las áreas rurales (Majerus, 1998). Este oscurecimiento gradual es resultado probable de la acción de la selección natural sobre una variación poligénica. Es plausible que el responsable sea la depredación selectiva de las formas menos crípticas, en zonas afectadas por contaminación atmosférica particulada. Si esta especulación resulta correcta, el reciente descenso en la contaminación debería conducir a una inversión de esta tendencia y los colores de fondo deberían volverse de nuevo más claros, y los diseńos más aparentes. Esto debería confirmarse mediante seguimiento en las próximas décadas. Los nuevos métodos de medida de la reflexión espectral de las superficies, y el desarrollo de cámaras digitales accesibles, debería permitir recoger y almacenar esa información sin tener que depender de especímenes de museos o de emulsiones fotográficas, que se decoloran con el tiempo.
Conclusión El melanismo industrial en los Lepidoptera, y en B. betularia en particular, es indudablemente más complejo de lo que usualmente se dice. La depredación diferencial por aves, junto con la migración, han sido los principales responsables del auge y caída de carbonaria. La opinión de que la selección no visual juega o ha jugado un papel importante en la evolución del melanismo industrial en Gran Bretańa carece de apoyo. La falta de concordancia entre las simulaciones y la frecuencia observada son, me parece, debidas más a una falta de comprensión del comportamiento de descanso de las polillas y a la interacción con sus depredadores que a factores selectivos aún no identificados. Esta opinión no es respaldada por todos los biólogos evolutivos y podría resultar incorrecta. No obstante, si ha de ser descartada, la investigación sobre B. betularia y otros casos debe continuar y ampliarse. En 1996 (New York Times, 12-XI-1996) Futuyma dijo sobre la historia de B. betularia: "No conocemos aún la historia completa, pero debería ser posible averiguar cuál es". Si las predicciones recientes sobre el declive de carbonaria (Mani & Majerus, 1993; Cook et al., en prensa) se cumplen, la apreciación de Futuyma podría ser excesivamente optimista pues quedan aproximadamente dos décadas antes de que carbonaria haya desaparecido del país en el que es mejor conocida.
Bibliografía Aldridge, D., Jones, C. W., Mahar, E. & Majerus, M. E. N., 1993. Differential habitat selection in polymorphic Lepidoptera in the Forest of Dean. Ent. Rec. J. Var., 105: 203-214. Asami, T. & Grant, B., 1995. Melanism has not evolved in Japanese Biston betularia (Geometridae). J. Lep. Soc., 49: 88-91. Barkman, J. J., 1969. The influence of air pollution on bryophytes and lichens. Air Pollution PUDOC, Wageningen: 197-209. Bates, J. W., Bell, J. N. B. & Farmer, A. M., 1990. Epiphyte recolonisation of oaks along a gradient of air pollution in south-east England, 1979-1990. Environ. Pollut., 68: 81-99. Bishop, J. A., 1972. An experimental study of the cline of industrial melanism in Biston betularia (L.) (Lepidoptera) between urban Liverpool and rural North Wales. J. Anim. Ecol., 41: 209-243. Bishop, J. A., Cook, L. M. & Muggleton, J., 1978. The response of two species of moths to industrialization in northwest England. I. Polymorphism for melanism. Phil. Trans. R. Soc. Lond. B, 281: 489-515. Boardman, M., Askew, R. R. & Cook, L. M., 1974. Experiments on resting site selection by nocturnal moths. J. Zool., 172: 343-355. Brakefield, P. M., 1987. Industrial melanism: do we have the answers? Trends Ecol. Evol., 2: 117-122. Brakefield, P. M., 1990. A decline of melanism in the peppered moth, Biston betularia in the Netherlands. Biol. J. Linn. Soc., 39: 327-334. Brakefield, P. M. & Liebert, T. G., 1990. The reliability of estimates of migration in the peppered moth Biston betularia and some implications of selection-migration models. Biol. J. Linn. Soc., 39: 335-342. Burkhardt, D. & Maier, E., 1989. The spectral sensitivity of a passerine bird is highest in the UV. Naturwissenschaften, 76: 82-83. Chen, D. & Goldsmith, T. H., 1986. Four spectral classes of cone in the retinas of birds. J. Comp. Physiol. A, 159: 473-479. Clarke, C. A. & Sheppard, P. M., 1964. Genetic control of the melanic form insularia of the moth Biston betularia L. Nature, 202: 215-216. Conroy, B. A. & Bishop, J. A., 1980. Maintenance of the polymorphism for melanism in the moth Phigalia pilosaria in North Wales. Proc. Roy. Soc. Lond. B, 210: 285-298. Cook, L. M., Askew, R. R. & Bishop, J. A., 1970. Increasing frequency of the typical form of the peppered moth in Manchester. Nature, 227: 1155. Cook, L. M., Rigby, K. D. & Seaward, M. R. D., 1990. Melanic moths and changes in epiphytic vegetation in north-west England and north Wales. Biol. J. Linn. Soc., 39: 343-354. Cook, L. M., Dennis, R. L. H. & Mani, G. S., en prensa. Melanic morph frequency in the peppered moth in the Manchester area. Proc. R. Soc. Lond. B. Edleston, R. S., 1864. No title (first carbonaria melanic of moth Biston betularia). Entomologist, 2: 150. Emmerton, J. & Delius, J. D., 1980. Wavelength discrimination in the "visible" and UV spectrum by pigeons. J.Comp. Physiol. A, 141: 47-52. Fisher, R. A., 1930. The Genetical Theory of Natural Selection. Oxford University Press, Oxford, 272 pp. Ford, E. B., 1940. Polymorphism and taxonomy. En: J. S. Huxley (ed.) The New Systematics. Clarendon Press, Oxford. Fraiers, T., Boyles, T., Jones, C. W. & Majerus, M. E. N., 1994. Short distance form frequency changes across habitat boundaries. Brit. J. Ent. Nat. Hist., 7: 47-52. Grant, B. & Howlett, R. J., 1988. Background selection by the peppered moth (Biston betularia Linn.): individual differences. Biol. J. Linn. Soc., 33: 217-232. Grant, B., Owen, D. F. & Clarke, C. A., 1995. Decline of melanic moths. Nature, 373: 565. Grant, B. S., Owen, D. F. & Clarke, C. A., 1996. Parallel rise and fall of melanic peppered moths in America and Britain. J. Hered., 87: 351-357. Grant, B. S., Cook, A. D., Clarke, C. A. & Owen, D. F., 1998. Geographic and temporal variation in the incidence of melanism in peppered moth populations in America and Britain. J. Hered., 89: 465-471. Guppy, C. S., 1986. The adaptive significance of alpine melanism in the butterfly Parnassius phoebus (Lepidoptera: Papilionidae). Oecologia, 70: 205-213. Haldane, J. B. S., 1924. A mathematical theory of natural and artificial selection. Trans. Camb. Phil. Soc., 23: 19-41. Hawksworth, D. L. & McManus, P. M., 1989. Lichen recolonisation in London under conditions of rapidly falling sulphur dioxide levels, and the concept of zone skipping. Bot. J. Linn. Soc., 100: 99-109. Heylaerts, F. J. M., 1870. Les macrolépidoptčres des environs de Bréda. Tijdschrift voor Entomologie, 13: 142-157. Hoffmeyer, S., 1949. De Danske Ugler. Universitetsforlaget I, Aarhus. Howlett, R. J. & Majerus, M. E. N., 1987. The understanding of industrial melanism in the peppered moth (Biston betularia) (Lepidoptera: Geometridae). Biol. J. Linn. Soc., 30: 31-44. Hudson, G. V., 1928. The Butterflies and Moths of New Zealand. Wellington, 386 pp. Jacobs, G. H., 1992. UV vision in vertebrates. Am. Zool., 32: 544-554. Jones, C. W., 1993. Habitat Selection in Polymorphic Lepidoptera. Ph. D. thesis, Cambridge University, 202 pp. Jones, C. W., Majerus, M. E. N. & Timmins, R., 1993. Differential habitat selection in polymorphic Lepidoptera. Entomologist, 112: 118-126. Kearns, P. W. E. & Majerus, M. E. N., 1987. Differential habitat selection in the Lepidoptera: a note on deciduous versus coniferous woodland habitats. Ent. Rec. J. Var., 99: 103-106. Kettlewell, H. B. D., 1955a. Selection experiments on industrial melanism in the Lepidoptera. Heredity, 9: 323-342. Kettlewell, H. B. D., 1955b. Recognition of appropriate backgrounds by the pale and black phases of the Lepidoptera. Nature, 175: 943-944. Kettlewell, H. B. D., 1956. Further selection experiments on industrial melanism in the Lepidoptera. Heredity, 10: 287-301. Kettlewell, H. B. D., 1958a. A survey of the frequencies of Biston betularia (L.) (Lepidoptera) and its melanic forms in Great Britain. Heredity, 12: 51-72. Kettlewell, H. B. D., 1958b. The importance of the micro-environment to evolutionary trends in the Lepidoptera. Entomologist, 91: 214-224. Kettlewell, H. B. D., 1973. The Evolution of Melanism. Clarendon Press, Oxford, 423 pp. Kettlewell, H. B. D. & Berry, R. J., 1969. Gene flow in a cline. Heredity, 24: 1-14. Kettlewell, H. B. D. & Conn, D. T., 1977. Further background-choice experiments on cryptic Lepidoptera. J. Zool., 181: 371-376. Kingsolver, J. G. & Wiernasz, D. C., 1991. Seasonal polyphenism in wing-melanin pattern and thermoregulatory adaptation in Pieris butterflies. Am. Nat., 137: 816- 830. Lees, D. R., 1981. Industrial melanism: Genetic adaptation of animals to air pollution. En: J. A. Bishop & L. M. Cook (eds.) Genetic Consequences of Man Made Change. Academic Press, Londres: 129-176. Lees, D. R. & Creed, E. R., 1977. The genetics of insularia forms of the peppered moth, Biston betularia. Heredity, 39: 67-73. Liebert, T. G. & Brakefield, P. M., 1987. Behavioural studies on the peppered moth Biston betularia and a discussion of the role of pollution and epiphytes in industrial melanism. Biol. J. Linn. Soc., 31: 129-150. Magnus, D. B. E., 1958. Experimental analysis of the Fritillary butterfly Argynnis paphia L. (Nymphalidae). Proc. 10th internat. Cong. Entomol., 2: 405-408. Majerus, M. E. N., 1981. The inheritance and maintenance of the melanic form nigrescens of Pachycnemia hippocastanaria (Lepidoptera: Geometridae). Ecol. Entomol., 6: 417-422. Majerus, M. E. N., 1982. Genetic control of two melanic forms of Panolis flammea (Lepidoptera: Noctuidae). Heredity, 49: 171-177. Majerus, M. E. N., 1989. Melanic polymorphism in the peppered moth Biston betularia and other Lepidoptera. J. Biol. Ed., 23: 267-284. Majerus, M. E. N., 1998. Melanism: Evolution in Action. Oxford University Press, Oxford, 338 pp. Majerus, M. E. N., Grigg, A., Jones, C. W., Salmon, F., Strathdee, A. & Dearnaley, N., 1994. Factors affecting habitat preferences in the Lepidoptera. Brit. J. Ent. Nat. Hist., 7: 129-137. Mani, G. S., 1980. A theoretical study of morph ratio clines with special reference to melanism in moths. Proc. R. Soc. Lond. B, 210: 299-316. Mani, G. S., 1982. A theoretical analysis of the morph frequency variation in the peppered moth over England and Wales. Biol. J. Linn. Soc., 17: 259-267. Mani, G. S. & Majerus, M. E. N., 1993. Peppered moth revisited: analysis of recent decreases in melanic frequency and predictions for the future. Biol. J. Linn. Soc., 48: 157-165. Mikkola, K., 1979. Resting site selection of Oliga and Biston moths (Lepidoptera: Noctuidae and Geometridae). Acta Entomol. Fenn., 45: 81-87. Mikkola, K., 1984. On the selective force acting in the industrial melanism of Biston and Oliga moths (Lepidoptera: Geometridae and Noctuidae). Biol. J. Linn. Soc., 21: 409-421. Mikkola, K., 1989. The first case of industrial melanism in the subarctic lepidopteran fauna, Xestia gelida f. inferna: new form of Noctuidae. Not. Entomol., 69: 1-3. Novak, I. & Spitzer, K., 1986. Industrial melanism in Biston betularia (Lepidoptera: Geometridae) in Czechoslovakia. Acta Entomol. Bohem., 83: 185-191. Owen, D. F., 1961. Industrial melanism in North American moths. Am. Nat., 95: 227-233. Partridge, J. C., 1989. The visual ecology of avian oil droplets. J. Comp. Physiol. A, 165: 415-426. Sargent, T. D., 1974. Melanism in moths of central Massachusetts (Noctuidae, Geometridae). J. Lep. Soc., 28: 145-152. Steward, R. C., 1977a. Multivariate analysis of variation within the insularia complex of the moth Biston betularia. Heredity, 39: 97-109. Steward, R. C., 1977b. Industrial melanism in the moths Diurnea fagella (Oecophoridae) and Allophyes oxyacanthae (Caradrinidae). J. Zool., 183: 47-62. Tutt, J. W., 1896. British Moths. George Routledge. Watt, W. B., 1968. Adaptive significance of pigment polymorphism in Colias butterflies, I: Variation of melanin in relation to thermoregulation. Evolution, 22: 437-458. West, B. K., 1977. Melanism in Biston (Lepidoptera: Geometridae) in the rural Appalachians. Heredity, 39: 75-81. West, B. K., 1992. Hydriomena impluviata D. & S. (Lep.: Geometridae): an extraordinarily rapid decline in melanism. Ent. Rec. J. Var., 104: 329. White, F. B., 1877. Melanochroism &c., in Lepidoptera. Entomologist, 10: 126-130. Whittle, P. D. J., Clarke, C. A., Sheppard, P. M. & Bishop, J. A., 1976. Further studies on the industrial melanic moth Biston betularia (L.) in the northwest of the British Isles. Proc. Roy. Soc. Lond. B, 194: 467-480. Wright, S., 1978. Evolution and the Genetics of Populations, Volume 4. Variability within and among Natural Populations. The University of Chicago Press, Chicago, IL, 580 pp. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
© 1999-2001 SEA, Sociedad Entomológica Aragonesa http://entomologia.rediris.es/sea - CV-e Comunidad Virtual de Entomología - http://entomologia.rediris.es - admin@entomologia.rediris.es |